Sepanta Laser Spadan
شرکت سپنتا لیزر اسپادان سهامی خاصSepanta Laser Spadan
شرکت سپنتا لیزر اسپادان سهامی خاصنظرسنجی
پیوندها
ارتباطات
- تماس با مدیر سایت نظرات و پیشنهادات خود را با مدیر سایت در میان گذارید
دستهها
ابر برجسب
تدریس آناتومی با شکوفه ساتری توموگرافی انسجام نوری لیزر شرکت سپنتا لیزر اسپادان رشته مهندسی اپتیک و لیزر تدریس فیزیک با شکوفه ساتری اپتیک پزشکی جزوه لیزر دندانپزشکی جزوه لیزر پزشکی گروه پژوهشی عطفان نژاد و ساتری جزوه لیزر گروه آموزشی مهندس شکوفه ساتری مهندس شکوفه ساتری بیوفوتونیک شکوفه ساتریبرگهها
جدیدترین یادداشتها
همه- حوزه های تحقیقاتی مهندس ساتری پژوهش حوزه های تحقیقاتی بیومکانیک مهندسی زیست مواد و احیا کننده مهندسی سلول و مولکولی تصویربرداری و بیوفوتونیک تجهیزات...
- اپتیک زیست پزشکی اپتیک زیست پزشکی رشته ای است که اصول اولیه برهمکنش بین نور و بافت های بیولوژیکی، سلول ها و مولکول ها را مطالعه می کند و...
- اپتیک زیست پزشکی فناوری های تصویربرداری نوری زیست پزشکی طراحی و ساخت دستگاه های نوری زیست پزشکی اپتیک بینایی، تصویربرداری و حس کردن چشم...
- بیوفوتونیک با شکوفه ساتری بیوفوتونیک بالینی و ترجمه ای توسعه و کاربرد تکنیک های نوری برای کاربردهای بالینی را پوشش می دهد. میکروسکوپ،...
- راهحلها و نوآوریها را از شرکتهای زیستپزشکی پیشرو در صنعت بیابید بزرگترین نمایشگاه اپتیک و بیوفوتونیک زیست پزشکی جهان است. جدیدترین فناوریها را از شرکتهای برتر ارائهدهنده راهحلهای...
- فناوری جدید هوش مصنوعی ممکن است به کشف عوامل درمانی برای اختلالات عصبی کمک کند یک گروه تحقیقاتی از دانشگاه ناگویا در ژاپن، هوش مصنوعی را برای تجزیه و تحلیل تصاویر سلولی ایجاد کرده است که از یادگیری...
- هدف محققان ایجاد یک الگوریتم هوش مصنوعی برای تشخیص زودهنگام اوتیسم در کودکان است آیا می توان از هوش مصنوعی برای کمک به تشخیص زودهنگام اختلال طیف اوتیسم استفاده کرد؟ این سوالی است که محققان دانشگاه...
- دستگاه جدید حسگر فیبر نوری، نشانگرهای زیستی کلیدی را به دنبال آسیب مغزی تروماتیک نظارت می کند دستگاه جدید حسگر فیبر نوری، نشانگرهای زیستی کلیدی را به دنبال آسیب مغزی تروماتیک نظارت می کند نتایج "امیدبخش"...
- کارآزمایی برتری هوش مصنوعی را در ارزیابی عملکرد قلب نسبت به ارزیابی سونوگرافی نشان می دهد کارآزمایی برتری هوش مصنوعی را در ارزیابی عملکرد قلب نسبت به ارزیابی سونوگرافی نشان می دهد در بیمارانی که تحت ارزیابی...
- هوش مصنوعی در تشخیص عملکرد قلب نسبت به سونوگرافیک ها دقیق تر است هوش مصنوعی در تشخیص عملکرد قلب نسبت به سونوگرافیک ها دقیق تر است در اولین کارآزمایی بالینی تصادفی شده در نوع خود که...
نویسندگان
بایگانی
تقویم
شهریور 1401| ش | ی | د | س | چ | پ | ج |
| 1 | 2 | 3 | 4 | |||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| 12 | 13 | 14 | 15 | 16 | 17 | 18 |
| 19 | 20 | 21 | 22 | 23 | 24 | 25 |
| 26 | 27 | 28 | 29 | 30 | 31 |
جستجو
کاوشگرهای مولکولی برای متابولیسم سلولی
متابولیسم مغز بسیار پویا است و با سطح محلی فعالیت عصبی و در نتیجه نیازهای انرژی در نوسان است. مقادیر قابل توجهی ATP برای پشتیبانی از گرادیان های یونی که به عنوان باتری برای فعالیت الکتریکی عمل می کنند مورد نیاز است و همچنین انرژی برای پشتیبانی از انتشار و بازیافت انتقال دهنده های عصبی مورد نیاز است. این انرژی عمدتاً توسط متابولیسم گلوکز تأمین می شود، اگرچه سایر مولکول های سوخت نیز می توانند در این امر نقش داشته باشند. سلولهای مغزی منفرد همچنین میتوانند بهطور اساسی در نیازها و پاسخهای متابولیکی خود متفاوت باشند: نه تنها ممکن است بین کلاسهای مختلف نورونها تفاوتهایی وجود داشته باشد، بلکه سلولهای گلیال مختلف - آستروسیتها (مرکز قسمت 3.7) و الیگودندروسیتها - ممکن است نقشهای خاصی در متابولیسم انرژی ایفا کنند. .
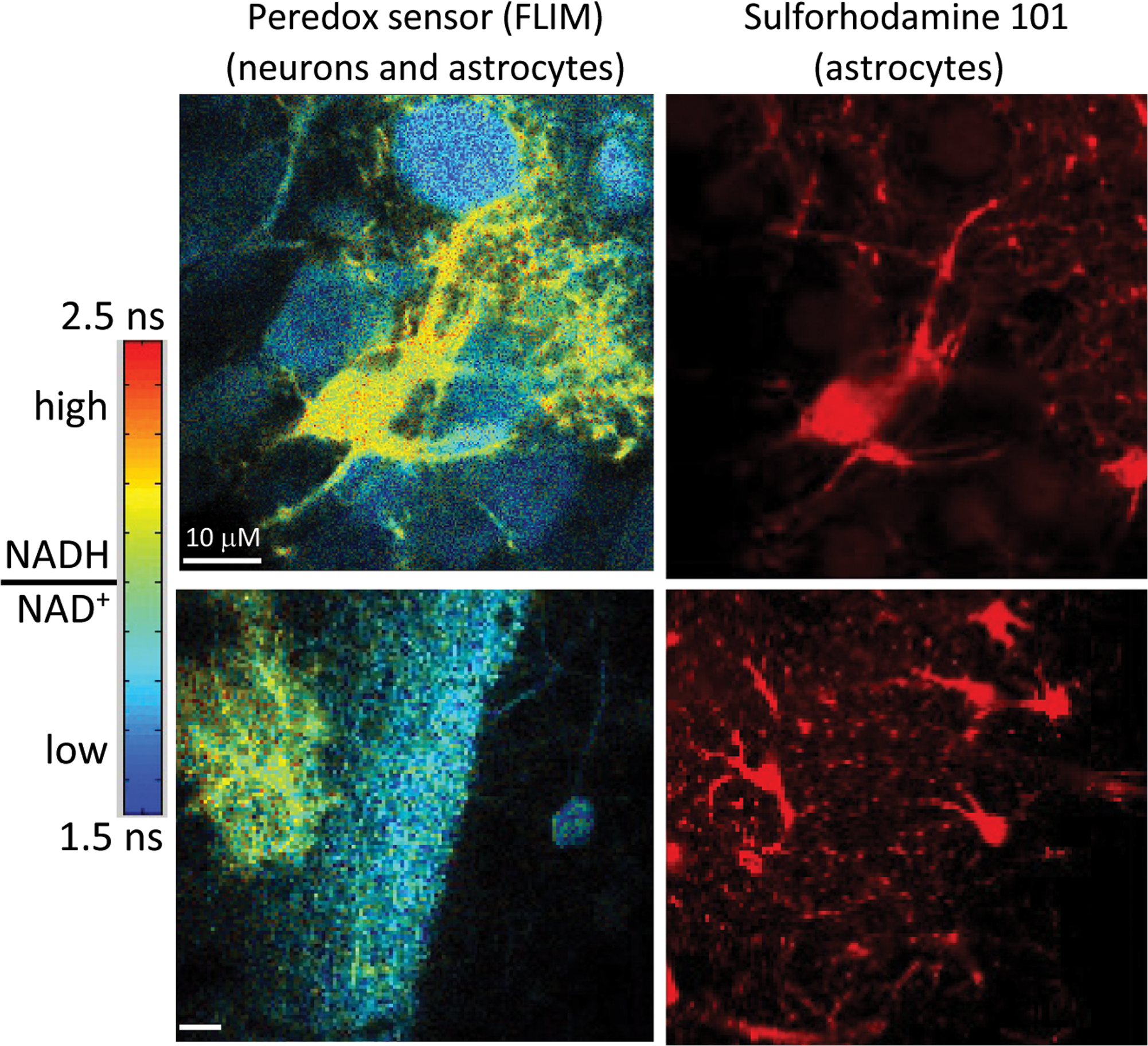
از آنجایی که پاسخهای متابولیکی سریع هستند (از چند ثانیه تا چند دقیقه) و میتوانند در سلولها و بخشهای درون سلولی موضعی شوند، تصویربرداری از حسگرهای زیستی فلورسنت میتواند رویکرد ایدهآلی برای بررسی نحوه عملکرد متابولیسم سلولی مغز باشد. حسگرهای زیستی رمزگذاری شده ژنتیکی برای گلوکز 227-230 و ATP؛ 231-233 برای کوفاکتورهای کلیدی حامل الکترون [خطای پردازش ریاضی] (شکل 13) و NADPH/NADP234-237 و برای واسطه های متابولیک پیرواتات238 و
لاکتات ایجاد شده اند. هم در شرایط in vitro و هم in vivo برای روشن کردن پاسخهای متابولیکی در نورونها و آستروسیتها تصویربرداری شدهاند، 240241 اگرچه برخی از ویژگیها کاملاً بحثبرانگیز هستند.